基因组不稳定性与 9-1-1 蛋白质复合体:
DNA中蕴藏着生物体的遗传信息,在细胞分裂过程中,DNA精确地复制并将其中的遗传信息传给下一代,DNA复制的高度忠实性,保证了生物遗传的稳定性。但是,生物体中细胞的DNA每时每刻都在遭受着损伤(图一),如果这些损伤得不到修复,细胞的正常生命活动就会受到破坏,由细胞构成的生命体就会产生突变,由此诱发一系列的遗传疾病,癌症的发生通常是由于基因的突变引起的。在大多数情况下,在人的生命中,基因组不稳定性(genomic instability)会最终导致足够的突变的积累并最终产生癌症(图二)。很多的DNA损伤都是自发产生的,甚至在一些情况下发生的频率很高,所以,在没有环境中DNA损伤试剂存在的情况下,DNA修复系统的缺陷也会引发很多的问题。
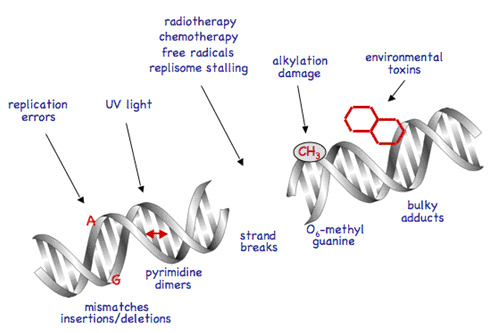
图一 DNA损伤
DNA损伤因素包括细胞内源和环境因素两方面。内源因素包括DNA复制错误、细胞代谢活动产生化学物质(如自由基、烷化剂)的攻击。外源因素包括紫外线、放射线、污染物、香烟燃烧剩余物和病毒等。
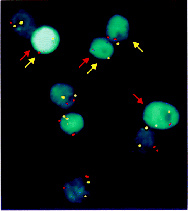
图二 基因组不稳定性
在正常细胞中有完善的DNA损伤修复和细胞周期监控点机制,维持基因组稳定。而恶性肿瘤细胞中DNA修复和细胞周期监控点机制有缺陷,基因组处于高度可变状态。基因组不稳定的细胞基因产生高频突变,改变细胞的各种特性,最终使细胞无控生长和恶性转移。图中细胞是从一个人结肠癌细胞分裂而来。红色的点为第三号染色体的着丝粒,黄色的点为第十二号染色体的着丝粒。正常细胞染色体成对,着丝粒也成对。但图中有的细胞只有一个红点,有点只有一个黄点,表明失去了着丝粒或染色体(Lengauer C et al., Nature,1998)。
DNA修复系统的缺陷的另一个问题是我们就不会从亲代那里得到遗传,从而物种的延续和生存就会出现问题。但是,DNA的复制又不能绝对的精确,否则就不能使物种进化以适应不断变化的环境。细胞内的多重DNA修复系统就是这两项工作的监控系统。一个有趣的现象是,所有的生物大分子中,只有DNA在受损后会进行修复,其它的大分子在受损后都只是被新的分子直接取代,这也从另一侧面印证了DNA在生物体中至关重要的地位。
造成DNA损伤的因素根据来源不同分为两类,第一类为细胞内源性损伤因素,包括以下几种:DNA复制错误;自发损伤包括碱基互变异构、碱基脱氨(C→U、A→I)和碱基丢失等;氧化代谢副产物如活性氧物质(Reactive oxygen species,ROS)的攻击等。第二类为环境中的损伤因素,包括辐射(含紫外线、X射线);化学致癌物(氧化脱氨,烷化剂或代谢活化物如苯并芘、黄曲霉素等产生碱基加合物)。
在漫长的进化过程中,生物体中进化出一套精密且有效的防御系统以应对随时发生的损伤,据统计,有一百多种基因编码的蛋白参与了DNA损伤修复。这套系统就像一个组织有序的行动小组,忙碌有序地工作以保护着生物体中基因组的稳定性。这些对DNA损伤的响应包括:(1)去除DNA损伤并且恢复DNA双链的连续性(2)激活细胞周期监控点,使细胞周期进程停滞以高效修复受损DNA (3)维持基因组稳定性,预防疾病发生。DNA损伤修复和细胞周期监控点是维持基因组稳定性的两个关键机制。恶性肿瘤细胞的DNA损伤修复和细胞周期调控过程均存在缺陷,因而基因组高度不稳定。
9-1-1蛋白质复合体是由Rad9、Hus1和Rad1构成的环状三聚体(图三)。9-1-1复合体可套在DNA双链上滑动。Rad9、Hus1和Rad1在生物进化上高度保守。人的Rad9基因可以部分代替裂殖酵母rad9基因的功能。9-1-1复合体在DNA损伤修复和细胞周期监控点机制中均发挥关键作用,是维持基因组稳定的重要因子。我们的研究显示,在小鼠身上敲除Rad9或Rad1均导致肿瘤易感(图四)。DNA损伤包括碱基水平、核苷酸水平、单链断裂、双连断裂、链内和链间交联等。生物进化出不同的修复途经来修复各类DNA损伤。我们及别的几个实验室的研究表明,9-1-1复合体参与多种DNA损伤的修复。
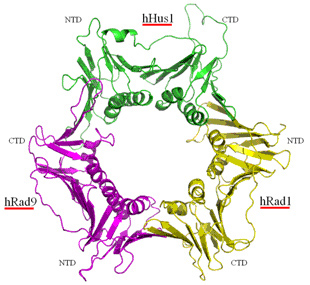
图三 9-1-1蛋白质复合体
9-1-1蛋白质复合体有Rad9、Hus1和Rad1构成。内径约30埃,可套在双链DNA上滑动 (Xu M et al, JBC, 2009)。
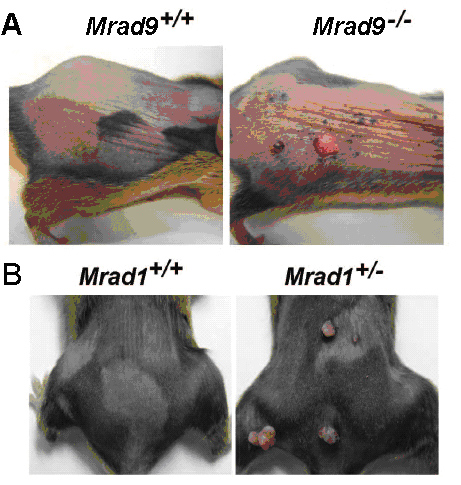
图四 Rad9或Rad1基因敲除导致小鼠肿瘤易感
在化学诱导剂作用下,纯合剔除Rad9或杂合剔出Rad1基因小鼠易感乳头瘤,而野生型小鼠则对肿瘤发生有抗性(Hu Z et al., Cancer Res. 2008; Han L et al., Molecular Cancer 2010)
尽管目前已知9-1-1复合体三维晶体结构和参与细胞周期监控点和几种DNA损伤修复的分子途径,但仍然不知为何9-1-1复合体能够参与众多不同的分子途径。我们实验室正利用分子生物学、细胞生物学、生物化学和基因敲除动物模型等多种研究手段解析这一9-1-1复合体功能的核心问题。我们同时积极研究肿瘤病人发病与Rad9、Hus1和Rad1缺陷的关系,希望能发现新的肿瘤诊断标记。
蛋白质人工进化:
过去数十年分子生物学、分子遗传学和细胞生物学的发展,尤其是近十多年发展起来的各类组学和生物信息学积累了大量生命过程的分子和细胞基本信息和运行机理。但是这些海量的知识并未衍生出一个类似于机械、电子或化工这样市场规模的工业行业。咎其原因,已有的各种生物学方法的叠加和直接利用别的已有工业行业的方法不能有效实施 “应用目标-设计-制造-组装-优化-产品”产品开发过程。居于这一认识,美国、欧洲和日本等国纷纷成立合成生物学中心、生物工程系,甚至生物工程学院,研究适用于生物工业的标准化部件、设计方法、优化原理等,为迎接生物产业的诞生做准备(图五)。
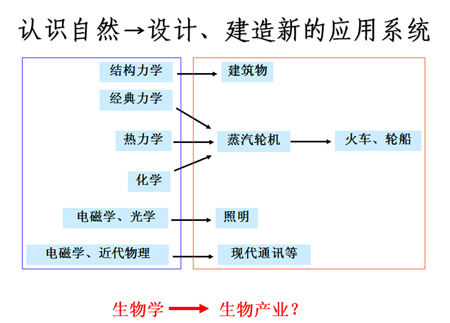
图五 迎接新兴生物产业
传统的生物工程技之所以没有催生一个像建筑业、机械制造、电子产业或化工产业一样的行业,其关键问题是生物过程更复杂、丰富和精密。不能采取现有简单的设计、制造、组装和优化技术。需要大力发展新的生物器件设计、制造和系统组装技术,同时发展生物器件和系统的优化技术。在机械或电子行业中,从设计部件和系统到组装成有功能的系统可以在短时间内完成。而目前如此设计、制造、组装出来一个新的生物系统需要非常长的时间,且多数组装的系统都是没有功能的或功能极其底下的。需要大力发展更精准的理性设计技术和更有效优化技术(如人工进化技术)。
在自然界,一个功能基因或蛋白质的发生和完善少则需要数百万年,多则需要数千万年甚至上亿年。最初通过基因扩增、融合、突变等过程形成新的基因。这些基因往往是没有功能或功能极其低下的,通过长期不断突变和环境压力的选择才衍生出具有精巧功能的基因和相应蛋白质。构象变化和精细识别是蛋白质区别于自然界和目前人造的别的微观物质的两个重要特性。这两个特性生物系统高效性的重要基础。既然不能通过简单的设计来获得这样精巧的分子,我们能否通过蛋白质设计、快速突变和高速功能筛选来创造这样东西呢?美国德克萨斯大学奥斯汀分校的Brent L. Iverson教授和George Georgiou教授实验室通过高速人工进化获得了依赖于位点硫酸化酪氨酸的蛋白质水解酶(Varadarajan N et al. Angew. Chem. 2008)。这类酶在自然界不存在。日本横滨庆应义塾大学的Hiroshi Yanagawa教授通过人工进化构建成β-内酰胺酶与绿色荧光蛋白融合蛋白(L-lactamase-GFP)。当该蛋白与β-内酰胺酶抑制蛋白结合时会诱导L-lactamase-GFP的构象变化并发出绿色荧光(图六)。该实验的重要性在于,在实验室条件下也能创造出具有构象变化的新蛋白质来。事实上,人工进化目前已成为疾病治疗抗体优化的关键手段。
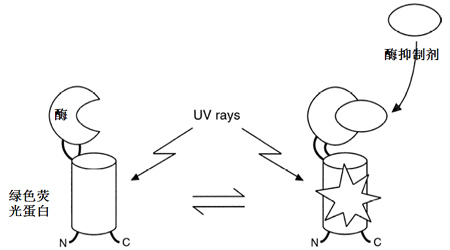
图六 可调绿色荧光蛋白质
Yanaganwa教授实验室用人工进化创造了新的绿色荧光蛋白质,用人工进化成功引入构象变化机制(Doi N et al., FEBS letter, 1999)。
自然界有数百万至上亿年的时间产生、完善一个具有功能的蛋白质,而属于人类的生物工业则没有这么长的时间。所以建立基因的快速突变和蛋白质功能的高速筛选方法十分关键。我们实验室最近成功建立两个新的高速蛋白质人工进化平台。一个是细菌蛋白展示和分选平台(Qiu J et al., Applied Microbiology and Biotechnology, 2010)。细菌平台可用于发现和分离与疾病细胞结合的新抗体和相应的疾病细胞靶抗原。采用同样的细菌平台对发现的抗体进行人工从进化,改进抗体的性能。这些抗体中可以开发出疾病诊断和治疗性抗体。另一个是人细胞蛋白展示分选平台。我们成功利用该平台快速人工进化了治疗自身免疫疾病的抗体。这两个人工进化平台可用于新酶、荧光蛋白质和别的功能蛋白创制和功能优化。
